 + 972547771177
+ 972547771177Некоторые основные факты о раке
 Раковые клетки утратили нормальные регуляторные механизмы, которые контролируют рост и размножение клеток.
Раковые клетки утратили нормальные регуляторные механизмы, которые контролируют рост и размножение клеток.- Раковые клетки потеряли способность дифференцироваться (что означает специализацию).
- Злокачественные раковые клетки проникают в новые ткани, чтобы создать вторичные опухоли, процесс, известный как метастазирование.
- Химические вещества, вызывающие рак, называются мутагенами.
- Рак может быть вызван химическими веществами, образом жизни (курением) и вирусами.
- Гены, которые связаны с причиной рака, называются онкогенами. Гены, которые меняются при мутации, называются протоонкогенами.
Признаки онкологии
- Самодостаточность сигналов роста (например, посредством активации онкогена H-Ras).
- Нечувствительность к сигналам, ингибирующим рост (анти-рост) (потеря супрессора ретинобластомы).
- Уклонение от запрограммированной гибели клеток (апоптоз) (продуцирует факторы выживания IGF).
- Безграничный репликативный потенциал (включая теломеразу).
- Длительный ангиогенез (продуцирует индуктор VEGF).
- Тканевая инвазия и метастазирование (инактивировать Е-кадгерин).
- Инактивация систем, которые регулируют в ответ на повреждение ДНК.
 Фазы клеточного цикла
Фазы клеточного цикла
- Фаза G1 (разрыв 1): клетка увеличивается в размерах и готовится скопировать свою ДНК в ответ на различные факторы роста.
- S фаза (синтез): репликация ДНК, копирование хромосомы.
- Фаза G2 (разрыв 2): подготовка к делению клеток. Проверка скопированной ДНК и восстановление поврежденной копии.
- M фаза (митоз): формирование митотического веретена и разделение на две отдельные клетки (деление клеток).
Контроль прогрессии клеточного цикла CDKs
- Прогрессирование через клеточный цикл контролируется циклин-зависимыми киназами (CDK).
- Связывание циклина со связанными с ним киназами запускает клеточный цикл в другую фазу.
- Присутствуют ингибирующие белки, которые могут модифицировать действие циклинов. К ним относятся р15 и р16, которые блокируют активность комплекса циклин D-CDK. Другим регулятором является р21, который контролируется белком-супрессором опухоли р53.
- Чрезмерно активные циклины или CDKs были связаны со многими опухолями. Чрезмерное производство циклинов или CDKs или недостаточное производство ингибиторов CDK приводит к нарушению нормальной регуляции клеточного цикла.
Смерть клетки
Некроз - это неконтролируемая (патологическая) гибель клеток. Есть много причин некроза, включая травмы, инфекцию, рак, инфаркт, токсины и воспаление.
Апоптоз - это запрограммированная гибель клеток. Он используется организмами для контроля количества клеток и размер ткани.
Способы апоптоза
Два пути активации: 1) на плазматической мембране через внешние лиганды при связывании с рецептором смерти или 2) через митохондриальный путь.
Связывание внешних лигандов, таких как рецептор фактора некроза опухоли (TNFα), с рецепторами Fas суперсемейства TNF индуцирует олигомеризацию рецептора и образование сигнального комплекса, индуцирующего гибель. Этот комплекс рекрутирует через молекулу-адаптер FADD (Fas-ассоциированный домен смерти) несколько молекул Pro-caspase-8, что приводит к активации caspase-8, что в итоге приводит к активации caspase-3.
В митохондриальном пути высвобождение апоптогенных факторов, таких как цитохром с, Apaf-1, каспаза-9-содержащий апоптосомный комплекс и белки-ингибиторы апопоза, запускают активацию каспазы-3
Существуют связи между двумя путями. Например, каспаза-8 приводит к расщеплению Bid, белка семейства Bcl-2, который перемещается в митохондрии для высвобождения цитохрома с.
Регуляторы апоптоза
- Семейство факторов Bcl-2 регулирует активацию каспазы либо отрицательно (например, Bcl-2, Bcl-XL, MCL1), либо положительно (например, Bcl-XS, Bax, BAD, BAK, BID).
- Ингибиторы апоптоза белков (IAP) замедляют апоптоз.
- Восходящими модуляторами являются онкогены, такие как c-myc, который активирует апоптоз таким образом, который важен для лечения рака.
- Опухолевый супрессор р53 индуцирует апоптоз при определенных обстоятельствах.
Теломеры
- Раковые клетки часто называют бессмертными, поскольку, нет предела тому, как часто они могут делиться.
- Время жизни нормальных клеток ограничено 50-60 делениями клеток. Это регулируется теломерами. Теломеры находятся на 3'-конце хромосом. После каждой репликации теряется около 50-100 пар оснований.
- В какой-то момент теломеры не эффективны, и ДНК становится нестабильной, что ограничивает репликацию. Раковые клетки обладают ферментом, называемым теломеразой, который поддерживает длину теломер и тем самым позволяет больше репликации ДНК.
Вторжение в ткани
- При злокачественных раковых клетках раковые клетки отрываются от первичного опухолевого участка, проникают в кровеносный или лимфатический сосуд, образуя участки метастазирования.
- Обычно клетки придерживаются только похожих клеток. Сигналы на поверхности клетки передаются через молекулы клеточной адгезии (например, кадгерины). Более того, клетки соединяются друг с другом путем установки их на внеклеточный матрикс (ЭМ).
- Адгезия к ЭМ включает молекулы, называемые интегринами.
- Металлопротеиназа белкового матрикса разрушает внеклеточный матрикс и поэтому важна для того, чтобы покинуть участок первичной опухоли и прикрепиться к вторичному участку.
- Если нераковая клетка отделяется от внеклеточного матрикса, она прекращает расти и запускается апоптоз.
- В метастазированных клетках молекулы клеточной адгезии отсутствуют, так что они могут покинуть участок первичной опухоли.
Развитие кровеносных сосудов
- Опухоли - быстро растущие ткани, которые должны иметь хорошее кровоснабжение.
- Ангиогенез относится к образованию новых кровеносных сосудов.
- Опухолевые клетки высвобождают факторы роста, такие как фактор роста эндотелия сосудов (VEGF) или фактор роста фибропластов (FGF-2), что приводит к прорастанию и расширению существующих капилляров.
- В здоровых тканях восстановление поврежденных тканей контролируется ингибиторами ангиогенеза, такими как ангиостатин и тромбоспондин.
- Кровеносные сосуды, возникающие в результате ангиогенеза, являются аномальными в том смысле, что они дезорганизованы по структуре и имеют утечки.
- Эти клетки отображают интегрины на своей поверхности, чтобы защитить новообразованные клетки от апоптоза.
- Прежде чем может начаться ангиогенез, необходимо разрушить базальную мембрану вокруг кровеносного сосуда (проводится с помощью матричных металлопротеиназ (ММР)).
Внутреннее подавление опухоли: р53
- В ответ на повреждение ДНК, активацию онкогена или другие вредные события индуцируется ген-супрессор опухоли p53.
- Различные киназы фосфорилируют р53, которые помогают стабилизировать его. Активированный р53 приводит к связыванию ДНК и активации транскрипции.
- MDM2 служит для подавления p53, что, в свою очередь, регулируется p14 ARF.
- р53 запускает остановку клеточного цикла в нетрансформированных клетках с помощью регуляторов клеточного цикла, таких как CDKs.
- Он также запускает апоптоз в трансформированных клетках через Bax.
- В большинстве опухолевых клеток р53 мутирует и инактивируется.
Лекарства, напрямую взаимодействующие с ДНК
- Алкилирующие агенты.
- Содержат высокоэлектрофильные группы.
- Образуют ковалентные связи с нуклеофильными группами в ДНК.
- Присоединяют N-1 и N-3 аденина и N-3 цитозина, и в частности N-7 оснований гуанина.
- Предотвращают репликацию и транскрипцию.
- Полезны в качестве противоопухолевых средств.
- Токсические побочные эффекты.
Нитрозомочевины
- Ломустин и кармустин являются жирорастворимыми и могут проникать через гематоэнцефалический барьер.
- Препараты разлагаются с образованием алкилирующих и карбамоилирующих.
- Образовавшийся изоцианат реагирует с группами NH3 лизина, инактивируя ферменты репарации ДНК.
- Алкилирующий агент реагирует сначала с O-6 гуанина, а затем с N-3 цитозина другой цепи.
Pt-алкилирующие агенты
- Связываются с ДНК в регионах, богатых гуаниновыми единицами.
- Запрещают транскрипцию.
- В растворе Cl-лиганды обмениваются на воду, что приводит к положительно заряженным лигандам, которые связываются с ДНК (с N-7 или O-6 соседних гуаниновых групп).
- Приводит к локализованному раскручиванию ДНК.
Антиметаболиты
- Ингибиторы рибонуклеотидредуктазы.
- Фермент превращает рибонуклеотиддифосфаты в дезоксирибонуклеотиддифосфаты, ингибируемые гидроксикарбамидом.
- Ингибиторы аденозин-деаминазы, например Пентостатин.
Противораковая терапия на основе гормонов
Стероидные гормоны связываются с ядерными рецепторами и действуют как факторы транскрипции.
Если для рака требуется определенный гормон, можно назначить гормон, вызывающий противоположный эффект.
Используемые: глюкокортикоиды (гормоны, участвующие в биосинтезе глюкозы, например, преднизолон), эстрогены, прогестины (например, ацетат медроксипрогестерона), аналоги гормонов, высвобождающих лютеинизирующий гормон (LHRH).
Препараты, действующие на структурные белки
Митоз - это упорядоченная серия событий, в которых идентичные копии генома перемещаются в отдельные места в делящейся клетке. Митотическая связь очень важна для этого события. Нити в митотическом веретене образованы из микротрубочек. Микротрубочки представляют собой элементы цитоскелета, присутствующие во всех эукариотических клетках. Они состоят из α- и β-субъединиц. Как образование (полимеризация), так и разрушение (деполимеризация) микротрубочек важны для правильного выполнения деления клеток. Лекарственные средства, препятствующие полимеризации / деполимеризации микротрубочек, влияют на митоз, вызывают остановку клеточного цикла и вызывают апоптоз.
Ингибиторы полимеризации тубулина
- Алкалоиды из Мадагаскарского барвинка.
- Таксол собирают с коры тисовых деревьев.
- Связывается с β-субъединицей тубулина и ускоряет полимеризацию.
- Полученные микротрубочки стабилизируются, ингибируя деполимеризацию.
- Клеточный цикл останавливается на стадии G2 / M.
- Приготовлен полусинтетически.
- Нельзя принимать внутрь.
- Вызывает множественную лекарственную устойчивость (субстрат для п-гликопротеина).
- Эфотилоны являются бактериальными метаболитами. Они не являются субстратом для P-GP.
Сигнальные пути, важные для раковой опухоли
Члены семейства Ras-белков принадлежат к GTPases и участвуют в передаче сигналов внутри клеток. Когда Ras «включен», он впоследствии включает другие белки, которые включают гены, участвующие в росте, дифференцировке и выживании клеток. В результате мутации в генах ras могут привести к выработке постоянно активированных белков Ras. Поскольку эти сигналы приводят к росту и делению клеток, сверхактивная передача сигналов Ras может в конечном итоге привести к раку.
p53 / HDM2: подавитель опухоли. р53 имеет множество механизмов противораковой функции и играет роль в апоптозе, стабильности генома и ингибировании ангиогенеза. Это модулируется MDM2.
PIK3: фосфатидилинозитол-3-киназа. PI3Ks представляют собой семейство ферментов-преобразователей сигнала, способных фосфорилировать 3-гидроксил инозитольного кольца фосфатидилинозитола. PI 3-киназы были связаны с клеточным ростом, пролиферацией, дифференцировкой, подвижностью, выживанием и внутриклеточным транспортом. Класс IA PI 3-киназа p110α мутирует во многих формах рака. Активность PI 3-киназы вносит значительный вклад в клеточную трансформацию и развитие рака.
AKT: протеинкиназа B (PKB, Akt) представляет собой серин-треонин-специфическую протеинкиназу, которая играет ключевую роль во многих клеточных процессах, таких как метаболизм глюкозы, апоптоз, пролиферация клеток, транскрипция и миграция клеток.
MAPK: Митоген-активируемые протеинкиназы (MAPK) - это протеинкиназы, специфичные для аминокислот серина, треонина и тирозина. Они регулируют функции клеток, включая пролиферацию, экспрессию генов, дифференцировку, митоз, выживание клеток и апоптоз.
Ингибиторы фарнезилтрансферазы
Сигнальный белок RAS участвует в раке. Мутации в RAS обнаружены в 30% раковых клеток. Мутант RAS является конститутивно активным.
Передача сигналов RAS требует связывания RAS с внутренней мембраной. Это делается путем добавления углеродной цепи фарнезилтрансферазой.
Ингибиторы тирозинкиназы
Низкомолекулярные ингибиторы тирозинкиназы (или TKI) - общие названия заканчиваются на «-nib». Обычно принимаются орально. Побочные эффекты варьируются в зависимости от того, какие ферменты они ингибируют (какова их цель). Некоторые из них эффективны против рака, устойчивого к большинству предыдущих методов лечения.
| Общее название | Бренд | Вид рака |
|---|---|---|
| Иматиниб | Gleevec | Лейкоз, рак желудка |
| Дасатиниб | Sprycel | Лейкоз, ОМЛ |
| Нилотиниб | Tasigna | Лейкоз |
| Гефитиниб | Iressa | Рак легких |
| Ерлотиниб | Tarceva | Рак легких, поджелудочной |
| Лапатиниб | Tykerb | Рак груди |
| Сорафениб | Nexavar | Рак почки, печени |
| Санитиниб | Sutent | Рак почки |
Ингибиторы факторов роста: Нацеливание на рецепторы EGF, рецептор TK.
Сверхэкспрессия измененных рецепторов эпидермального фактора роста приводит к образованию онкогена (EGFR, семейство ERBB).
Ингибитор тирозинкиназы Абельсона (BCR-ABL).
- Киназа BCR-ABL является единственным онкогеном, ответственным за редкий рак крови.
- Ингибирование аутофосфорилирования BCR-ABL по Gleevec.
- Обработка трансформированных BCR-ABL клеточных линий Gleevec приводит к зависимому от дозы снижению роста опухоли.
- Противоопухолевый эффект специфичен для клеток, экспрессирующих BCR-ABL.
- Gleevec повторно активирует апоптоз в клетках BCR-ABL.
Ингибиторы ангиогенеза, нацеленные на рецептор VEGF, белок тирозин.
- Повышенные уровни факторов роста фибропластов (FGF) и рецептора сосудистого эндотелиального фактора роста (VEGF) связаны с ангиогенезом.
- VEGF регулируется несколькими цитокинами, например, трансформирующий фактор роста (TGF-β), эпидермальный фактор роста (EGF) и тромбоцитарный фактор роста (PDGF).
- Ингибиторы в основном нацелены на связывание АТР с другими мишенями для киназы: тромбоцитарные факторы роста (PDGF-R), митоген-активируемые протеинкиназы (MAPK), рецептор фактора роста инсулина 1 (IGF-1R), протеинкиназа B (PKB), c-Src тирозинкиназа, инозиттрифосфаткиназа (IP3K).
Ингибиторы циклинзависимых киназ, S / T киназы
- CDK важны для контроля клеточного цикла (в основном на G1 / G2 в зависимости, например, от повреждения ДНК).
- Ser / Thr киназы
- Они активируются циклинами и ингибируются ингибиторами циклинзависимой киназы.
Ингибиторы матриксных металлопротеиназ
- ММР являются цинк-зависимыми ферментами (протеазами).
- Чрезвычайно разрушительные ферменты, участвующие в ремоделировании внеклеточного матрикса или соединительной ткани.
- ММР включают каллагеназы, желатиназы, стромелизины и мембранный тип (МТ).
- Они подавляют ангиогенез.
- Коллагеназа расщепляется между глицином и изолейцином.
Ингибиторы протеасомы
Является единицей для разложения поврежденных или неправильно свернутых белков, но также разлагает белок, участвующий в регуляции. Белки, отмеченные для деградации, помечены убиквитином. Ингибирование протеасомы приводит к накоплению регуляторных белков, таких как промотор апоптоза Bax. Накопление регуляторных белков приводит к клеточному кризису и запускает апоптоз.
Лечение рака антителом
 Уничтожение опухолевых клеток с использованием моноклональных антител (mAbs) может быть результатом прямого действия антитела (например, посредством блокады рецепторов), иммуноопосредованных механизмов уничтожения клеток, доставки полезной нагрузки и специфического воздействия антитела на сосудистую сеть опухоли и стромы.
Уничтожение опухолевых клеток с использованием моноклональных антител (mAbs) может быть результатом прямого действия антитела (например, посредством блокады рецепторов), иммуноопосредованных механизмов уничтожения клеток, доставки полезной нагрузки и специфического воздействия антитела на сосудистую сеть опухоли и стромы.
Опухолевые антигены, которые были успешно нацелены, включают рецептор эпидермального фактора роста (EGFR), ERBB2, фактор роста эндотелия сосудов (VEGF), цитотоксический антиген 4, ассоциированный с Т-лимфоцитами (CTLA4), CD20, CD30 и CD52.
Серологические, геномные, протеомные и биоинформационные базы данных также использовались для идентификации антигенов и рецепторов, которые сверхэкспрессируются в популяциях опухолевых клеток или которые связаны с мутациями гена, определяемыми как движущие силы пролиферации раковых клеток (опухолевые маркеры).
Основной целью клинической оценки mAb было определение токсичности и терапевтической эффективности антитела в отдельности или в качестве системы доставки для радиоизотопов или других токсических агентов. Также важно оценить его специфичность in vivo путем определения его биораспределения у пациентов и оценить соотношение поглощения антител в опухоли по сравнению с нормальными тканями. Двенадцать антител (2012 г.) получили одобрение FDA США для лечения различных солидных опухолей и гематологических злокачественных новообразований, и большое количество дополнительных терапевтических антител в настоящее время тестируется на ранних и поздних стадиях клинических испытаний.
Номенклатура моноклональных антител
Последний слог всегда – маб.
- Рядом с последним слогом
▪ -u- человек (100%). Панитумумаб.
▪ -зу - гуманизированный (95%): Трастузумаб.
▪ -си - химерный (65%): Ритуксимаб.
▪ -о - мышь, -а - крыса, -е - хомяк, -и - примат: Тоситумомаб.
- Предыдущий слог
▪ -tu (m) - для общей опухоли [-ma (r) - молочная железа, -pr (o) - простата, -co (l) - толстая кишка и т. д.)
▪ -ci (r) - для кровообращения: Бевацизумаб.
Цели антител
| Мишени антиангиогенных mAbs | VEGF | Bevacizumab | Опухоли сосудистой сети |
|---|---|---|---|
| VEGFR | IM-2C6 and CDP791 | Опухоли сосудистой сети | |
| Integrin αVβ3 | Etaracizumab | Солидные опухоли | |
| Integrin α5β1 | Volociximab | Опухоли сосудистой сети | |
| Сигналы роста и дифференцировки | EGFR | Цетуксимаб, панитумумаб, нимотузумаб и 806 | Опухоли глиомы, легких, молочной железы, толстой кишки, головы и шеи |
| ERBB2 | Трастузумаб и пертузумаб | Опухоли груди, толстой кишки, легких, яичников и простаты | |
| ERBB3 | MM-121 | Опухоли груди, толстой кишки, легких, яичников и простаты | |
| MET | AMG 102, METMAB и SCH 900105 | Опухоли груди, яичников и легких | |
| IGF1R | AVE1642, IMC -A12, MK-0646, R1507 и CP 751871 | Глиома, рак легких, молочной железы, головы и шеи, рак простаты и щитовидки | |
| EPHA3 | KB004 и IIIA4 | Опухоли легких, почек и толстой кишки, меланома, глиома и рак крови | |
| TRAILR1 | Мапатумумаб (HGS-ETR1) | Опухоли толстой кишки, легких и поджелудочной и рак крови | |
| TRAILR2 | HGS-ETR2 и CS -1008 | Опухоли толстой кишки, легких и поджелудочной и рак крови | |
| RANKL | Деносумаб | Рак простаты и метастазы в кости | |
| Антигены стромального и внеклеточного матрикса | FAP | Сибротузумаб и F19 | Опухоли толстой кишки, груди, легких, поджелудочной, головы и шеи |
| Tenascin | 81C6 | Глиома, опухоль молочной железы и простаты |
| Антигенная категория | Примеры антигенов | Примеры терапевтических mAbs | Типы опухолей, экспрессирующих антиген |
|---|---|---|---|
| Антигены кроветворной дифференцировки | CD20 | ритуксимаб | Неходжкинская лимфома |
| Ибритумомаб, тиуксетан и тозитумомаб | Лимфомы | ||
| CD30 | Брентуксимаб ведотин | Ходжкинская лимфома | |
| CD33 | Гемтузумаб озогамицин | Острый миелогенный лейкоз | |
| CD52 | Алемтузумаб | Хронический лимфолейкоз | |
| Гликопротеины | EpCAM | IGN101 and адекатумумаб | Эпителиальные опухоли |
| CEA | Лабетуцумаб | Опухоли груди, толстой кишки и легких | |
| gpA33 | huA33 | Колоректальный рак | |
| Mucins | Пемтумомаб и ореговомаб | Опухоли груди, толстой кишки, легких и яичников | |
| TAG-72 | CC49 (минретумомаб) | Опухоли груди, толстой кишки и рак легких | |
| CAIX | cG250 | Карцинома | |
| PSMA | J591 | Рак простаты | |
| Folate-binding protein | MOv18 and MORAb-003 (фарлетуцумаб) | Рак яичников | |
| Гликолипиды | Gangliosides (such as GD2, GD3 and GM2) | 3F8, ch14.18 and KW-2871 | Нейроэктодермальные и эпителиальные опухоли |
| Углеводы | Le | hu3S193 and IgN311 | Опухоли груди, толстой кишки, легких и простаты |
Т-клеточный иммунный ответ
 Т-клеточная вызванная иммунная реакция включает образование антигена, Т-клеточного рецептора и MHC (основного комплекса гистосовместимости). Она представляет фрагменты белков и пептидов, обработанных клеткой (нет необходимости подвергаться воздействию поверхности). Затем запускает иммунный ответ (производство цитокинов, факторов лизирования клеток и т. д.)
Т-клеточная вызванная иммунная реакция включает образование антигена, Т-клеточного рецептора и MHC (основного комплекса гистосовместимости). Она представляет фрагменты белков и пептидов, обработанных клеткой (нет необходимости подвергаться воздействию поверхности). Затем запускает иммунный ответ (производство цитокинов, факторов лизирования клеток и т. д.)
Антитела / Т-клеточный ответ
После проникновения антигены поглощаются (в основном дендритными клетками), а белки разлагаются внутри. Фрагменты представлены на поверхности в сочетании с основным комплексом гистосовместимости (MHC). Несамостоятельные белки или капсульные полисахариды активируют B-клетки через B-клеточный рецептор (BCR). Исходные антитела - это преимущественно низкоаффинный иммуноглобулин-M (IgM) abs. Повторное воздействие вызывает более быстрый иммунный ответ. Антитела преимущественно класса IgG или IgA. У них вообще намного более высокое родство.
Аналогично, в 10 000 раз больше цитотоксических Т-клеток.
Рецептором на В-клетках является мембраносвязанный иммуноглобулин (BCR), который может распознавать целые белки. В противоположность этому рецепторы Т-клеток (TCR) распознают только небольшие пептиды. В- и С-клетки распознают разные эпитопы.
В наивной (неоспоримой) популяции В- и Т-клеток присутствуют клетки, способные распознавать все эпитопы.
При заражении антигеном быстро размножаются только соответствующие В-клетки, которые могут распознавать антиген (клональная экспансия). Точно так же Т-клетки подвергаются аналогичной активации.
Повышается аффинность связывания антител (созревание аффинности). Во время этого процесса происходит много мутаций в гипервариабельных петлях (соматическая гипермутация). Во время этого процесса может происходить переключение классов антител
Иммунотерапия рака: блокада иммунных контрольных точек
Опухоли приводят к множеству мутаций в генных продуктах, которые могут быть распознаны как лишние и запускают иммунный ответ для их очистки. Иммунные контрольные точки относятся к ингибирующим путям иммунной системы, которые имеют решающее значение для поддержания самостоятельной толерантности и модуляции продолжительности и амплитуды, физиологических иммунных реакций в периферических тканях, чтобы минимизировать повреждение коллатеральных тканей. Опухоли неправильно используют иммунную контрольную точку, чтобы избежать чистки иммунной системы, в частности, чтобы избежать специфичных для опухолевого антигена Т-клеточных ответов. Иммунные контрольные точки часто инициируются взаимодействиями лиганд-рецептор, и они могут блокироваться антителами или модулироваться рекомбинантными формами из лигандов или рецепторов. Эти антитела не нацелены на опухолевую клетку, но нацелены на молекулы, участвующие в регуляции Т-клеток, солдат иммунной системы. Цель иммунной контрольно-пропускной терапии состоит не в том, чтобы активировать иммунную систему для атаки определенных целей на опухолевые клетки, а в том, чтобы устранить ингибирующие пути, которые блокируют эффективные противоопухолевые Т-клеточные ответы.
Блокада иммунных контрольно-пропускных пунктов (II)
Важным иммунным контрольным пунктом является цитотоксический антиген 4, ассоциированный с Т-лимфоцитами (CTLA4), который подавляет активацию Т-клеток. Запрограммированный белок 1 гибели клеток (PD1) ограничивает эффекторную функцию Т-клеток в тканях. Повышая регуляцию лигандов для PD1, опухолевые клетки блокируют противоопухолевый иммунный ответ в микроокружении опухоли. Напротив, разрушение опухоли, вызванное иммунотерапией, часто задерживается или даже предшествует периоду видимого роста опухоли. Только часть пациентов реагирует на блокаду иммунных контрольных точек. В этих случаях микроокружение опухоли может быть не иммуногенным. Комбинированная терапия может затем создать иммуногенную микросреду, которая реагирует на иммунную контрольную терапию. Многие из этих методов лечения ограничены токсичностью.
Иммунная контрольная точка, опосредуемая цитотоксическим Т-лимфоцит-ассоциированным антигеном 4 (CTLA4), индуцируется в Т-клетках во время их первоначального ответа на антиген. Уровень индукции CTLA4 зависит от амплитуды исходной передачи сигналов, опосредованной рецептором Т-клеток (TCR). Высокоаффинные лиганды индуцируют более высокие уровни CTLA4, что снижает амплитуду первоначального ответа. Наивные и Т-клетки памяти экспрессируют высокие уровни CD28 клеточной поверхности, но не экспрессируют CTLA4 на своей поверхности. Вместо этого CTLA4 секвестируется во внутриклеточных везикулах. После того, как TCR запускается при обнаружении антигена, CTLA4 транспортируется на клеточную поверхность. Чем сильнее стимуляция через TCR (и CD28), тем больше количество CTLA4, которое откладывается на поверхности Т-клеток. Следовательно, CTLA4 функционирует как демпфер сигнала, чтобы поддерживать постоянный уровень активации Т-клеток в условиях широко варьирующихся концентраций и сродства лиганда к TCR.
Основная роль пути запрограммированного клеточного гибели белка 1 (PD1) заключается не в начальной стадии активации Т-клеток, а скорее в регуляции воспалительных реакций в тканях эффекторными Т-клетками, распознающими антиген в периферических тканях. Активированные Т-клетки активируют PD1 и продолжают экспрессировать его в тканях. Воспалительные сигналы в тканях индуцируют экспрессию лигандов PD1, которые подавляют активность Т-клеток и, таким образом, ограничивают повреждение коллатеральных тканей в ответ на инфекцию микроорганизмов в этой ткани. Лучше всего охарактеризованным сигналом для индукции PD1-лиганда 1 (PDL1; также известным как B7-H1) является интерферон-γ (IFNγ), который преимущественно продуцируется клетками T-хелпера 1 (TH1)
Иммунотерапия рака II: реинжиниринг Т-клеток: CAR T-клетки
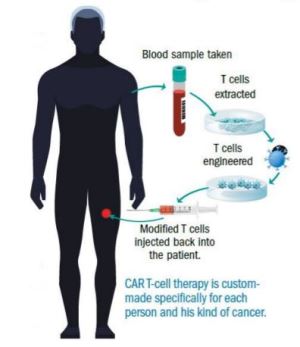 Врачи собирают Т-клетки пациента. Они помещают белок снаружи (цитотоксических) клеток. Этот белок является либо рецептором Т-клеток, либо рецептором химерного антигена (CAR). Затем сконструированные Т-клетки вводят обратно пациенту. Добавленный белок выполняет две роли: он направляет Т-клетку непосредственно к опухоли, и по прибытии он запускает боевую силу Т-клетки, чтобы атаковать раковые клетки.
Врачи собирают Т-клетки пациента. Они помещают белок снаружи (цитотоксических) клеток. Этот белок является либо рецептором Т-клеток, либо рецептором химерного антигена (CAR). Затем сконструированные Т-клетки вводят обратно пациенту. Добавленный белок выполняет две роли: он направляет Т-клетку непосредственно к опухоли, и по прибытии он запускает боевую силу Т-клетки, чтобы атаковать раковые клетки.
Т-клетки взаимодействуют через свои Т-клеточные рецепторы с белками основного комплекса гистосовместимости (MHC), которые представляют опухолевые антигены на поверхности опухолевых клеток. Т-клетки могут быть реинжинирированы для представления антителоподобных молекул (клеток CAR) на их поверхности. Антитела часто представляют собой одноцепочечные AB, направленные против антигенов опухолевых клеток.
 Онлайн видеоконсультация с ведущими израильскими специалистами :
Низкая стоимость, Кратчайшие сроки.
Подробно о преимуществах онлайн видеоконсультации.
Онлайн видеоконсультация с ведущими израильскими специалистами :
Низкая стоимость, Кратчайшие сроки.
Подробно о преимуществах онлайн видеоконсультации.




")





